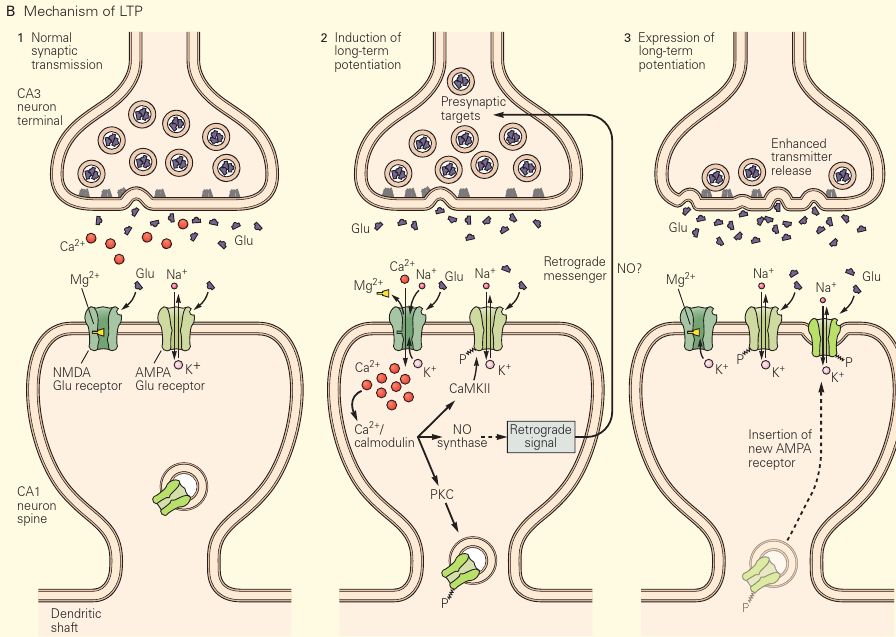
LTP
2022-04-19: reference:
LTP #
-
LTP can be descibed as something like: NMDAR activation in causes a persistent increase in synaptic transmission via modification of postsynaptic AMPARs, whether by:
- Increase in number
- Probability of being open
- Kinetics
- Single-channel conductance (γ)
- Modulation of AMPA receptor unitary conductance by synaptic activity: Here we show that the induction of LTP in the CA1 region of the hippocampus is often associated with an increase in single-channel conductance of AMPARs.
-
-
 ?
? -
Genes and signaling pathways involved in memory enhancement in mutant mice
-
- GluR1/GluR2 receptors are added to synapses during plasticity; this requires interactions between GluR1 and group I PDZ domain proteins (SAP-97). In contrast, GluR2/GluR3 receptors replace existing synaptic receptors continuously
- So, under things like Ser831 (induced by Insulin, fresh GluR1-containing AMPARs receptors are differentially modified before being recycled into a mature GluR3, with much less phosphorylation. (and then an association with GRIP and PICK, which GluR2 already had, but still)
-
Morphological changes in dendritic spines associated with long-term synaptic plasticity (2001)
- After LTP/induction of CREB, structural changes in Dendrites ensue, such as the formation of a mushroom shape (enlarging of the head + thickening & shortening of the spine)
Early-Late LTP #
- Gene transcription->Protein synthesis is thought to converge on PI3K/PKA/PKC/CAMKII->ERK.
- In contrast to some claims (at least on Wikipedia):
Long-lasting LTP requires neither repeated trains for its induction nor protein synthesis for its development (2012)
- But, perhaps some people by definition differentiate E-LTP and L-LTP by the presence of protein synthesis, making this ad hoc.
- A single high-frequency train could trigger an LTP lasting more than 8h that was not affected by either anisomycin or cycloheximide (two inhibitors of protein synthesis).
- Persistent LTP without triggered protein synthesis (2008): Effects lasting longer than about 2-3h are considered to require synthesis of new proteins.
-
EVIDENCE FOR CONSTITUTIVE PROTEIN SYNTHESIS IN HIPPOCAMPAL LTP STABILIZATION: Continuous protein synthesis application after LTP induction did not cause an observable effect on LTP stabilization.
- The idea that LTP is stabilized beyond its first few hours by proteins whose synthesis is triggered by the LTP-inducing high-frequency stimulation rests almost entirely on the observation from numerous studies that application of PSIs during the ‘‘critical’’ time window results in an LTP that is largely decayed within few hours.
- The findings reported by Fifkova et al. (1982) were problematic in term that early (4 min) but not late (90 min) Entorhinal Cortex spine morphological changes were prevented by administration of anisomycin in vivo.
- Structural basis of long-term potentiation in single dendritic spines (Matsuzaki 2004) dendritic spine enlargement merely 1 min after glutamate receptor stimulation.
- The findings reported by Fifkova et al. (1982) were problematic in term that early (4 min) but not late (90 min) Entorhinal Cortex spine morphological changes were prevented by administration of anisomycin in vivo.
- The idea that LTP is stabilized beyond its first few hours by proteins whose synthesis is triggered by the LTP-inducing high-frequency stimulation rests almost entirely on the observation from numerous studies that application of PSIs during the ‘‘critical’’ time window results in an LTP that is largely decayed within few hours.
Calcium control hypothesis #
-
See also with AMPARs: AMPAR
-
LTD stimulus is characterized as low, but persistent Ca2+ activity, while LTP stimulus is characterized by high, quicky reversed Ca2+(i.e. 15 min 900 pulses at 1 Hz vs. 2x1 sec 100 pulses at 100 Hz) . This Ca2+ is therefore mediated by CaMKII in a differential manner as explored in @Autonomous CaMKII mediates both LTP and LTD using a mechanism for differential substrate site selection (Coultrap et al. 2014).
- I guess to understand it better I can consider it: fast AND sufficient vs low OR prolonged.
-
Are these weak+persisent and high+quick always mutually inclusive? Or is a high Ca2+ influx regulated somehow to be quick? Gotta learn how this is mediated. I think ‘high’ and ‘persistent’ could be an oxymoron, since insertion of new AMPA would make it effectively low? Something like that. This is what this stuff is all about in the first place!
- Reminds me of the signal-noise dynamic in @Memantine a NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system - too little activation is bad, too much is even worse (Parsons et al. 2007) so clearly this is some kind of well-recognized thing for LTP. I think that paper has a section on kinetics I could check out.
- I mean, AMPA is very much characterized by fast (in)activation, while NMDA is much slower. Clearly, this is the key. Also relevant (though maybe not too significant) is how GluR1 homomers allow Ca2+ influx which seems trippy. It’s all coming together here, though.
- Reminds me of the signal-noise dynamic in @Memantine a NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system - too little activation is bad, too much is even worse (Parsons et al. 2007) so clearly this is some kind of well-recognized thing for LTP. I think that paper has a section on kinetics I could check out.
-
Selective Induction of LTP and LTD by Postsynaptic Ca2+i Elevation
- When the results of Figs. 1 and 2 are considered together, they demonstrate that neither a difference in the amplitude of Ca2+i elevation nor a difference in duration is sufficient by itself to determine the response of synapses to this chemical signal.
- However, the principle does still hold: both forms of synaptic plasticity can be reliably and selectively induced by different patterns of Ca2+i elevation, without the need to invoke any other Ca2+-independent signaling or triggering process.
- All I want to know is WTF prolonged high means? More or less, LTP = amplitude, LTD = duration. I guess it would be 50/50.
- However, the principle does still hold: both forms of synaptic plasticity can be reliably and selectively induced by different patterns of Ca2+i elevation, without the need to invoke any other Ca2+-independent signaling or triggering process.
- The two cascades (Phosphatases/LTD vs. kinases/LTP) may interact and inhibit each other at one or more intermediate stages. Such a scheme of enzyme interactions resembles the multilayered and mutually inhibitory neural circuits involved in pattern recognition
Neural Networks for Pattern Recognition (Bishop 1995). The mechanisms of pattern recognition in “enzyme circuits” may be similar to those of neural circuits.
- Holy shiet… that’ a book, btw. 11,000 citations.
- When the results of Figs. 1 and 2 are considered together, they demonstrate that neither a difference in the amplitude of Ca2+i elevation nor a difference in duration is sufficient by itself to determine the response of synapses to this chemical signal.
-
A calcium-based plasticity model predicts long-term potentiation and depression in the neocortex
- This is known as the calcium control hypothesis, or the differential threshold hypothesis.
-
- DO I HEAR FOUR?: The synaptic tagging hypothesis points out that a strong activation (e.g., strong tetanization) of synapses potentiates those synapses by a dual mechanism—setting synapse-specific “tags” and triggering the synthesis of plasticity-related products (PRPs). In addition, other synapses that form a synaptic tag but express a transient synaptic potentiation (early LTP) due to a weak tetanization can capture and utilize the previously formed PRPs to stabilize their potentiation.
- The STC hypothesis highlights the fact that synaptic efficacy is shaped by neural activity occurring on a multitude of time scales: Making memories last: The synaptic tagging and capture hypothesis
- DO I HEAR FOUR?: The synaptic tagging hypothesis points out that a strong activation (e.g., strong tetanization) of synapses potentiates those synapses by a dual mechanism—setting synapse-specific “tags” and triggering the synthesis of plasticity-related products (PRPs). In addition, other synapses that form a synaptic tag but express a transient synaptic potentiation (early LTP) due to a weak tetanization can capture and utilize the previously formed PRPs to stabilize their potentiation.
-
Probably speaks on it: Sensitivity of CaM kinase II to the frequency of Ca2+ oscillations